|


 n
order to better understand the universe in which we
live, MedTab introduces the "chicken" from a far plant
called "Earth." During a recent visit to Earth, Doctor
Treslegs, a member of the Alien Abduction Society (AAS),
was able to sit in on a "developmental biology"
conference, undetected. According to the information he
gathered it appears as though us Puppeteers develop
within the womb in an orderly fashion through the
joining of certain elements. n
order to better understand the universe in which we
live, MedTab introduces the "chicken" from a far plant
called "Earth." During a recent visit to Earth, Doctor
Treslegs, a member of the Alien Abduction Society (AAS),
was able to sit in on a "developmental biology"
conference, undetected. According to the information he
gathered it appears as though us Puppeteers develop
within the womb in an orderly fashion through the
joining of certain elements.
This article, specifically, has to do
with a process the humans call, induction. Induction can
be thought of as a signaling between cells in which one
cell (or a group of cells) sends a message to responder
cells in order to stimulate (or inhibit) certain
qualities. These qualities include, be are not limited
to: stimulation or inhibition of mitosis within the
responding cells; stimulation of the responder cells to
secrete signaling molecules; and stimulation of the
responding cells to restrict their potency and become
more specified (toward their final phenotype). As you
will see, humans have found, for example, that lenses
within the eye are derived from the ectoderm. Through a
series of inductive cues, the ectoderm invaginates and
becomes the lens. In this case, the tissue which sends
the message to the ectoderm (and thus inducing it) is
the optic vesicle of the eye.
Can all tissue types respond to all
inducing signals?
It turns out that the answer to this
question is NO. Using the example of the lens from
above, it turns out that if the optic vesicle is
transplanted to other regions of the body containing
ectoderm, the lens will fail to develop because the
ectoderm is not "competent" to respond to the inducing
signal of the optic vesicle. Thus, both the inducing
tissue and the responder tissue, and their spatial
relationship to each other, are incredibly important for
normal development. We invite you to explore our website
and learn about how induction aids in the formation of
many body parts.
Below we begin by briefly touching on the
topics of fertilization, cleavage, gastrulation,
neurulation, and organogenesis.
Fertilization
Fertilization is the combination of two haploid gametes
to form a single diploid cell. In sexually reproducing
species a male sperm combines with a female ovum (egg)
to create a zygote. Once the diploid zygote cell is
created it can divide and specialize to give rise to the
individual organism.
The reaction between a sperm and an egg is a complicated
and regulated mechanism. For starters, the sperm has to
have the ability to enter the egg and transfer its
genetic information. To enter the egg sperm have an
enzymatic layer, the acrosome. The enzymes released by
the acrosome digest the zona pellucida of the egg (Tosney
2005). Once through the zona pellucida the sperm can
fuse with the egg membrane to create one membrane (Fox
1998).
At this point regulation is very important; the egg
cannot allow two sperm fertilize it. If two sperm
fertilize an egg a condition, called polyspermy, occurs
and the cell dies. To prevent polyspermy the
fertilization pathway has two key regulatory mechanisms.
The first way to prevent polyspermy is a mechanism
called the fast block (Tosney 2005). In the fast block
mechanism the fertilized egg changes its electrical
potential. The depolarization change in electrical
potential prevents additional sperm from binding.
However, this is not a permanent solution to preventing
polyspermy (Fox 1998). So, the egg also uses a method
called the slow block.
Upon sperm entering the egg, a rush of calcium ions
trigger the cortical granules release enzymes that
initiate the slow block response (Fox 1998). The
cortical granules fuse with the plasma membrane of the
egg and enzymatic activity dissolve protein posts that
hold the vitelline and plasma membrane together. This
reaction causes water to rush into the egg which expands
the vitelline envelope creating a fertilization
membrane. The fertilization member prevents additional
sperm from fertilizing the egg (Tosney 2005).
For information on Puppeteer fertilization, please
consult the “Sexes” page.
Cleavage
Cleavage immediately follows fertilization, and is a
series of extremely rapid mitotic divisions in which the
enormous volume of zygote cytoplasm is divided into
numerous smaller cells, eventually forming a sphere-like
structure known as the blastula (Gilbert, 2003).
The mammalian cleavage process has been noted to contain
many unique properties. This process is among the
slowest in the animal kingdom, as it generally takes
about 12-24 hours. The orientation of mammalian
blastomeres is another unique aspect, as it undergoes
the cleavage pattern known as rotational cleavage
(Gilbert, 2003).
Mammalian cleavage is also unique in that the
blastomeres do not all divide at the same time, and thus
mammalian embryos do not increase exponentially, but
often contain an odd numbers of cells. Unlike other
non-mammalian development, the switch from mother to
zygotic genome becomes activated during early cleavage,
allowing for the zygote to produce the necessary
proteins for continued cleavage and development to occur
(Gilbert, 2003).
However, the most crucial difference to mammalian
cleavage is the phenomenon of compaction. Compaction
involves the increased expression of cell adhesion
proteins, such as E-cadherin, inducing the blastomeres
to become adherent and spread on each other to form a
compact ball of cells (Tosney, 2005). This tightly
packed arrangement is stabilized by tight junctions that
form between the outside cells, while leaving gap
junctions that allow for small molecules and ions to
pass between them (Gilbert, 2003).
Image of mammalian zygote undergoing compaction

©2004
Gynaecworld.com
The 8-cell embryo continues to divide to produce a
16-cell morula, which subsequently divide to become the
trophoblast cells, which produce the extra-embryonic
structures, while the inner cell mass will give rise to
the embryo and its associated yolk sac, allantois, and
amnion (Gilbert, 2003). The process of cleavage appears
to be similar in the Pierson’s Puppeteer.
Image of trophoblast and inner cell mass formation
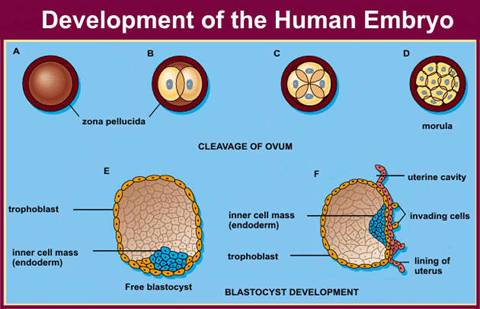
©2000-2004 Becomehealthynow.com
(Click on Image to view Original Source)
Gastrulation
After cleavage has ceased, the blastomeres undergo
dramatic movement where they change their positions
relative to one another. This series of extensive cell
rearrangement is called gastrulation and is very
critical for the developing embryo. The end of
gastrulation occurs by the formation of a gastrula and
the three germ layers, ectoderm, mesoderm and endoderm.
These layers specify all other parts of the developing
embryo.
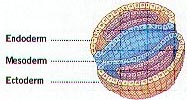
© 2005 by Jerry Johnson
In the bird and the mammal development the primitive
streak forms, which defines the axes of the embryo and
starts in the posterior of the animal and moves towards
the anterior. Cells ingress along the primitive streak,
forming the three major layers of the embryo mentioned
above. At the head fold, the node develops and moves
from the anterior to the posterior, laying down the
notochord. This notochord induces pattern and neural
tissue creating a gradient of maturity leaving the
anterior structures more developed than the posterior
structures.
This is a SEM of the primitive streak in a chick embryo.

Used with permission from © K. Tosney
Once this process of gastrualation occurs, the
anterior/posterior axis is specified by HOX gene
expression. In the alien development gastrulation
occurs in much of the same way; however there is rapid
convergence with the different expression of HOX genes.
Most importantly, in the case of the alien gastrulation
begins in the posterior end of the embryo and since the
HOX gene code is reversed, the posterior end is in the
top of the neck for the developing alien.
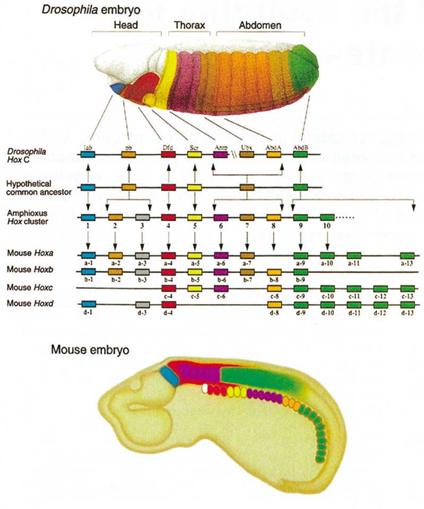
© 2002 by Henry Gee
(Click on Image to view Original Source)
Neurulation
In mammals there are two mechanisms by which the neural
tissue of the body is formed. The anterior undergoes
primary neurulation while the posterior undergoes
secondary neurulation. In primary neurulation the
epithelium thickens and many processes occur (i.e.
convergent extension, spread of the ectoderm, anchorage
of the underlying cells, etc.) to cause the epithelium
to elevate and fold. there is then cell-cell
recognition, differential adhesion, and reorganization,
which allow closure to occur, giving rise to a neural
tube and an overlying ectoderm.
In secondary neurulation, which occurs in the trunk of
the presumptive spinal cord, a process known as
cavitation occurs, forming a hollow neural tube.
Differences with this mechanism from primary neurulation
results from differential gene expression.
Organogenesis
What is Organogenesis?
Organogenesis is the development of the organs and
tissues of an embryo. This process begins after
gastrulation when the three embryonic tissue layers
(endoderm, mesoderm, and ectoderm) develop. The
organogenesis of a mouse and of a Puppeteer are fairly
similar, differing mostly with respect to brain
organogenesis.

Formation of the Heart
The heart is one of the first structures to develop
during organogenesis. The heart is formed from
splanchnic mesoderm (the most ventral mesoderm) (Tosney).
The heart originally forms outside the body as a
thickened tube. It later develops its chambers and
ingresses into the body cavity (Tosney). The endoderm
regresses to form a place for the heart, and no endoderm
is in the heart (Tosney). A Puppeteer’s heart is one of
the first organs to form, as in the mouse. Their hearts
usually are larger, with the left ventricle
significantly larger than the mouse’s because the heart
must pump blood up two necks instead of one.
Otherwise, a Puppeteer’s heart forms the same way as the
mouse heart does.
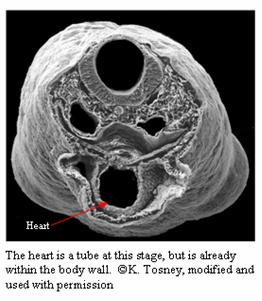
Formation of the Brain
The brain also begins development early, although it
continues to develop through gestation and even after
birth (Tosney). The brain’s neurons are “born” at
different times and migrate to the outermost layer of
the brain. Each layer migrates past the old layers to
the outermost portion of the brain; therefore the brain
has an “inside-out” organization (Tosney). There are
three general types of neurons: sensory, motor, and
commissural. Sensory neurons have axons in the brain,
spinal cord, muscles, and skin (Tosney). Motor neurons
have axons in muscles (Tosney). Commissural neurons
have axons that stay in the brain and spinal cord (Tosney).
Some of the axons in the body are myelinated (surrounded
by cells) to send signals more quickly. In the central
nervous system (brain and spinal cord), oligodendrocytes
ensheath the axon. In the peripheral nervous system
(nerves not in the brain or spinal cord), Schwann cells
ensheath the axon (Gilbert). A Puppeteer’s brain is
more specialized than a mouse brain. In the mouse, the
brain is right in the head, close to the eyes and
mouth. In a Puppeteer, the brain is located under the
main between the two necks’ connection to the rest of
the body. A Puppeteer’s brain, therefore, forms in the
inside-out organization, but the two brain hemispheres
grow away from each other, instead of next to each other
as in the mouse, in order to accommodate the optic and
other nerves that go to the eye, head, muscles, and
mouth.

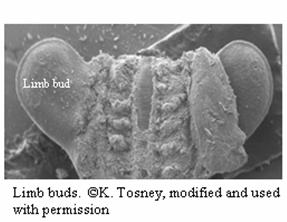
Formation of the Limbs
The limbs form from outgrowths of the ectoderm called
limb buds and their muscles come from somites
(developmental units that form during neurulation)
(Tosney). Mesenchyme from somites migrates to the
inside of the limbs and will form bones and muscles.
The Puppeteer embryo has three limb buds, two for the
from legs and one for the back leg. The necks and heads
do not form from limb buds, but rather from a splitting
of the region of the embryo anterior to the brain.
Formation of the Urinary System
The urinary system begins with the formation of the
kidney. Nephrogenic mesenchyme (of mesodermal origin)
near the posterior portion of the embryo develops into
the pronephros and the nephric duct (the primordial
kidney) (Tosney). The pronephros extends posteriorly
down the nephric duct, finally degenerating as its
replacement, the mesonephros, forms (Tosney). The
mesonephros will 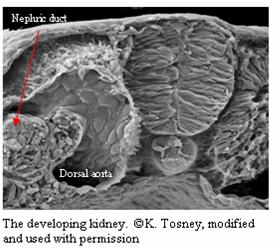 be replaced with the final kidney, the metanephros (Tosney). The dorsal aorta interacts with
the mesonephros to form the glomerulus, which is the
part of the kidney that has contact with many blood
vessels (Tosney). Waste leaves the blood and enters the
kidney. The metanephros will induce the metanephric
mesenchyme to form the ureters, which lead to the
bladder where urine is stored (Tosney). As the embryo
develops, the kidneys move anteriorly to their proper
location (Tosney). Puppeteers have urinary systems that
are fairly similar to a mouse urinary system. be replaced with the final kidney, the metanephros (Tosney). The dorsal aorta interacts with
the mesonephros to form the glomerulus, which is the
part of the kidney that has contact with many blood
vessels (Tosney). Waste leaves the blood and enters the
kidney. The metanephros will induce the metanephric
mesenchyme to form the ureters, which lead to the
bladder where urine is stored (Tosney). As the embryo
develops, the kidneys move anteriorly to their proper
location (Tosney). Puppeteers have urinary systems that
are fairly similar to a mouse urinary system.
Formation of the Digestive and Respiratory Systems
The organs of the digestive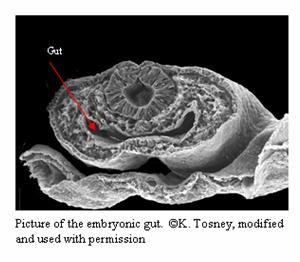 system and respiratory
system all branch off from the embryonic gut. The lungs
branch off the from the anterior portion of the gut;
while the liver, spleen, gall bladder, and pancreas
branch off from the middle part of the gut (Gilbert).
The intestines are just specialized portions of the gut
tube, as are the esophagus and stomach (Liem et al.).
The formation of mouse and Puppeteer digestive and
respiratory systems are the same, with the notable
exception that the Puppeteer has two esophagi that both
empty into the same stomach. system and respiratory
system all branch off from the embryonic gut. The lungs
branch off the from the anterior portion of the gut;
while the liver, spleen, gall bladder, and pancreas
branch off from the middle part of the gut (Gilbert).
The intestines are just specialized portions of the gut
tube, as are the esophagus and stomach (Liem et al.).
The formation of mouse and Puppeteer digestive and
respiratory systems are the same, with the notable
exception that the Puppeteer has two esophagi that both
empty into the same stomach.
Sources:
Fox, Richard. Lecture Notes Bio 112. Lander
University. <http://www.lander.edu/rsfox
/112devel.html>. 1998.
Gilbert, Scott F. Developmental Biology. 7th
ed. Sunderland: Sinauer Associates, Inc., 2003. p.
364-368
Liem, Karel F., et al. Functional Anatomy of the
Vertebrates: An Evolutionary Perspective. 3rd
ed. Brooks/Cole: Belmont, 2001.
Tosney, Kathryn W. “Heart and circulatory system.”
Biology 208: Embryology. Ann Arbor. 17 Nov. 2005.
Tosney, Kathryn W. “Limb Development.” Biology 208:
Embryology. Ann Arbor. 29 Nov. 2005.
Tosney, Kathryn W. “Neural Development.” Biology 208:
Embryology. Ann Arbor. 27 Oct. 2005.
Tosney, Kathryn W. “Urinary System.” Biology 208:
Embryology. Ann Arbor. 15 Nov. 2005.
Tosney, K. "Fertilization." Biology 208: Embryology.
Ann Arbor. 2005.
Tosney, Kathryn W. “Early Vertebrate Development.”
Biology 208: Embryology. Ann Arbor. September 29,
2005.
 |
