|
Claws/Hooves Development & Induction
Normal Development:
So much of development is due to epithelial-mesenchymal
interactions, where almost always the mesenchyme
provides the initial signal that induces the responding
tissue, the epithelial. Hoof and claw development is a
perfect example; they are formed because of a series of
reciprocal exchanges of inductive signals. In the case
of limb development it is the somatic mesoderm that is
responsible for the initial signal to the limb dermis.
There is a limb morphogenic field that is committed to
give rise to the limb and will do so upon receiving the
right induction signal. Tbx4 is a transcription factor
that is expressed in the hind limbs and in combination
with a growth factor, FGF, the limb out grows and forms
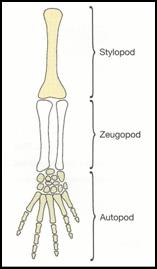
the hooves. As the limb grows outward, the stylopod
forms first, followed by the zeugopod and then the
autopod; this shows how the tissues are differentiated
in the proximal to distal order. Each phase of limb
development requires a particular expression of hox
genes. BMPs are expressed in the distal area of the
sclerotome and BMPs both induce apoptosis to form the
digits but also help differentiate mesenchyme cells into
cartilage. In normal development BMPs are inhibited so
that apoptosis can take place and digits and joints are
formed. Claws form similar in which a claw field
develops as an epithelial thickening on the dorsal side
of each digit. This epithelial thickening, or placode,
is the first sign of induction for an epithelial
appendage. Next to this placode a transverse groove
appears and forms a deep fold in the epidermal matrix
and the cells of this matrix produce a keratinized layer
that slides distally over the claw bed. (Hamrick 2001).
© 1999 by John Stear
(Click on Image to view Original Source)
How we differ:
Although much of our development appears to be very
similar to this so-called chick creature, we do differ
in some of the hooves and claw development. It appears
that our development is very similar for the stylopod
and zeugopod, but differs at the autopod region.
Whereas the chick embryo secretes noggin, a BMP
inhibitor, it is obvious that we lack this factor
because it is the increase expression of BPM that
converts the mesechyme of the digits to cartilage which
will later form hard keratin and form the hooves
(Gilbert 2003). Our specific claw pattern develops
through interactions with SHH which is expressed in the
region of the limb bud called the zone of polarizing
activity (ZPA), which is a region
of mesoderm located between the limb bud and the body
wall which affects the development of digits in the
limb. SHH induces a signal that affects the surrounding
hox genes and this epithelial-mesenchyme interaction is
what results in the claws of the hooves. We seem
to display a Hoxd-13 mutation, (as those people on earth
call it) which is why the claws on its hooves can fuse
together. Thereby, three claws form instead of the
normal five found in most vertebrates on earth. In
creatures on earth, this is considered a deformity known
as syndactyly, as seen below. This syndrome is
analogous to the our normal claw.

© 2005 by Jon A. Baskin
(Click on Image to view Original Source)


© 2005 Florida Museum of Natural History
© 1996-2003 Nesssus
(Click on Image to view Original Source)
(Click on Image to view Original Source)
The central digit becomes increasingly stronger while
the “side digits” become less important and are
virtually lost in us.
Sources:
Gilbert, Scott F. Development Biology. 7th
Editon. Massachusetts: Sinauer Associates, Inc., 2003.
Hamrick, Mark W. “Development and evolution of the
mammalian limb: adaptive diversification of nails,
hooves, and claws.” Evolution and Development. Volume 3,
Issue 5, Page 355. September-October 2001.
Tosney, K. Lecture Material. Bio 208, Fall 2004
 |
