|
Two Neck
Phenotype
Normal
Mammalian Neck Development:
In mammals, the neck
develops in coordination with the rest of the trunk. In
short, the primitive streak moves from the posterior to
the anterior of the embryo creating a grove (“streak”)
in the tissue. The streak signals the surrounding to
migrate inward, toward the center of the embryo. This
migration and convergence of the epiblast toward the
streak and eventually through it creates more
specialized tissue layers, such as endoderm and
mesenchymal mesoderm.
When the streak reaches its
anterior most limit it begins to move back from where it
came from (the posterior). At this point the leading
edge of the migrating cell mass is referred to as
Hensen’s Node (HN). HN’s main task is to lay down the
notochord as it moves toward the posterior of the embryo
which, in turn, creates a gradient of maturity with the
more mature area of the embryo lying toward the anterior
pole.
Once HN has passed and the
notochord has been laid, the ectoderm on both sides of
the midline begins to thicken and then elevate.
Following elevation these two cell populations hinge
toward the midline, meet, combine, and eventually
separate from the overlying ectoderm to create a hollow
neural tube separate from the overlying ectoderm it was
derived from. Additionally, neural crest cells appear at
the ectoderm-neural tube boarder, near the midline.
These cells are also derived from ectoderm. Following
their formation the overlying epidermis induces them to
express Slug and RhoB proteins by secreting BMP-4 and
BMP-7. This induction is highly important because this
causes the neural crest cells to lose their tight
junctions and and N-Cadherin expression, thus allowing
them to migrate. If Slug or RhoB protein expression is
suppressed or halted, the neural crest cells will fail
to migrate.
Next, the segmental plate
which lies laterally of the neural tube on both sides
and is composed of mesoderm begins to segment (or pinch
off) into individual somites at the rate of 1 pinched
off somite for every 90 minutes. This temporal clock is
thought to be regulated by the Lfng, Hes1, Hes7, Hey2
genes (Erol, et al., 2003).
Picture showing the
"pinching off" of somites.
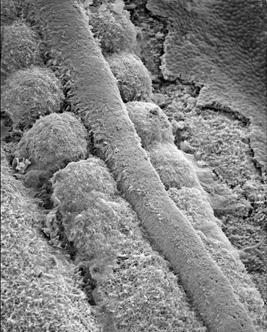
Used and modified with permission © K Tosney
Now that the neural tube,
somites, and various tissue layers are present, the
embryo is ready to continue forming its central nervous
system and begin forming its peripheral nervous system,
vertebra, and (more toward the posterior) viscera, ribs
and other anatomic features of the mammal (see
organogenesis).
The actual formation of the
neck in mammal, as us Peppeteers think of it, has mostly
to do with their neural tube, somites, and neural crest
cells. The somites, of which part will become vertebra,
not only are segmented themselves, but also lead to
segmentation of the migrating neural crest cells and
thus, the autonomic nervous system (ANS).
Neural Crest Migration:
After the somites have
formed they further differentiate into three main
tissues: dermatome, myotome, and sclerotome. If the
somite is yet to differentiate and form these three
tissues, the neural crest will not migrate from the
neural tube. It isn’t until the somites form the three
cell populations mentioned above that the neural crest
cells begin to migrate from their starting positions.
Once the neural crest cells sense sclerotome they begin
to migrate into it. This migration, however, does not
happen at all locations along the somites. Only the
anterior most pole of each somite allows for the
migration of neural crest cells. This is due to the
posterior inhibitory somite tissue that expresses ephrin
proteins which the neutral crest cells can recognize
with their eph receptors. Thus, the somites create a
segmental pattern along the anterior-posterior (AP)
axis. Furthermore, since the PNS is derived from neural
crest cells it exhibits a segmental pattern also.
Picture showing the
migration of neural crest cells.
 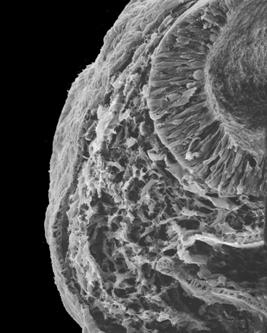
 Used
and modified with permission © K Tosney Used
and modified with permission © K Tosney
Somites:
To enable flexibility
within the neck (and back), the neck is composed of
vertebra. These so called vertebra are develop from
medial sclerotome in an interesting fashion. To form the
vertebra the medial sclerotome divides horizontally (in
a transecting fashion). Following this division the
anterior half of one joins with the posterior half of
the adjacent segment. Since somites are present on both
sides of the neural tube, the same process occurs on
both sides and, synchronously, fuse at the dorsal and
ventral midline surrounding the neural tube (Peck, et
al., 2003).
In order for the sclerotome
to form the vertebral column, Sonic hedgehog (SHH)
expression appears to be obligatory (Bonaventure, 2003).
The notochord contributes the SHH. Additionally,
different vertebral segments are coded for by Hox gene
expression (Bonaventure, 2003). Following cartilaginous
vertebra formation, the cartilage ossifies due to the
implantation of osteoblasts into the cartilage at the
ossification center. The osteoblasts secrete a calcium
salt matrix which mineralizes the bone.
________________________________________________________________________________________________
Puppeteer
Development
As described in the eyes
and brain sections, the neural tubes which the
Puppeteer’s necks develop from the diencephalon where
the formation of the optic vesicle causes the underlying
neural tissue to undergo rapid mitosis. So, neck
development in us is different than mammalian neck
development. Our neck’s neural tissue is not derived
from the tissue running parallel to the primitive streak
and Hensen’s Node. Instead, they are formed as described
above and migrate toward the anterior pole. During the
migration of the neural tubes, overlying ectoderm must
“stretch” toward the anterior to accommodate the
developing neural tubes. Additionally, the migration of
the neural tubes stimulates the mesenchymal mesoderm to
undergo mitosis. Thus, when the neural tubes reach their
desired height, the surrounding ectoderm and mesoderm
have compensated and are sitting in the same position as
we see in mammals.
Following the migration of
the neural tube and subsequent compensation of the
ectoderm and mesoderm, the somites begin to form
bilaterally, as they do in mammal. At about the same
time the neural crest cells are expelled from the neural
tube due to induction of these cells by the overlying
ectoderm (the gene/cue is yet to be determined). At this
point the migration of the neural crest cells begin in
the segmented pattern seen in mammals, only after they
have been induced by the myotome to express RhoB and
Slug proteins. In Puppeteers this segmental pattern is
due to the expression of Ephrin proteins on the
posterior sclerotome. In Puppeteers, the migration of
neural crest cells in predominately responsible for the
formation of sensory neurons within the necks. Once the
neural crest cells have migrated the sclerotome begins
to form the vertebra and, like in mammal, the sclerotome
splits and the bottom of one segment fuses with the top
of another. Following this fusion, the vertebra
encompass the neural tube and fuse medially. Your necks
are now formed. Since the necks contain vertebra they
are highly flexible, yet sturdy, and pack enough
strength to protect the afferent optical signals
entering the brain. Afterall, a Puppeteer who can’t see
is no Puppeteer at all!
Sources:
Bonaventure J. “Skeletal
development in human: a model for the study of
developmental genes.”
Atlas of Genetics and Cytogenetics in Oncology and
Haematology. University Hospital, 2003.
Erol B. Kusumi
K. Lou J.
Dormans
JP. “Etiology
of Congenital Scoliosis.” UPOJ
(15) Pages 37-42. 2002.
Peck WW. Hesselink JR. Barkovich JA.
“Pediatric
Spinal Anomalies.”
University of California – San Diego, Dept. of
Neuroradiology. 2003.
Online at: http://spinwarp.ucsd.edu/NeuroWeb/Text/sp-140.htm
Tosney K.W. “Neural
Crest.” Biology 208: Embryology, Ann Arbor. 2005.
Tosney K.W. “Somites”
Biology 208: Embryology, Ann Arbor. 2005.
 |
